- Фагоцит
-
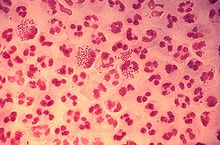
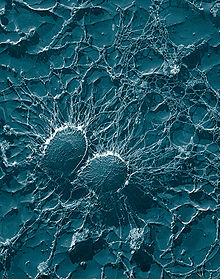
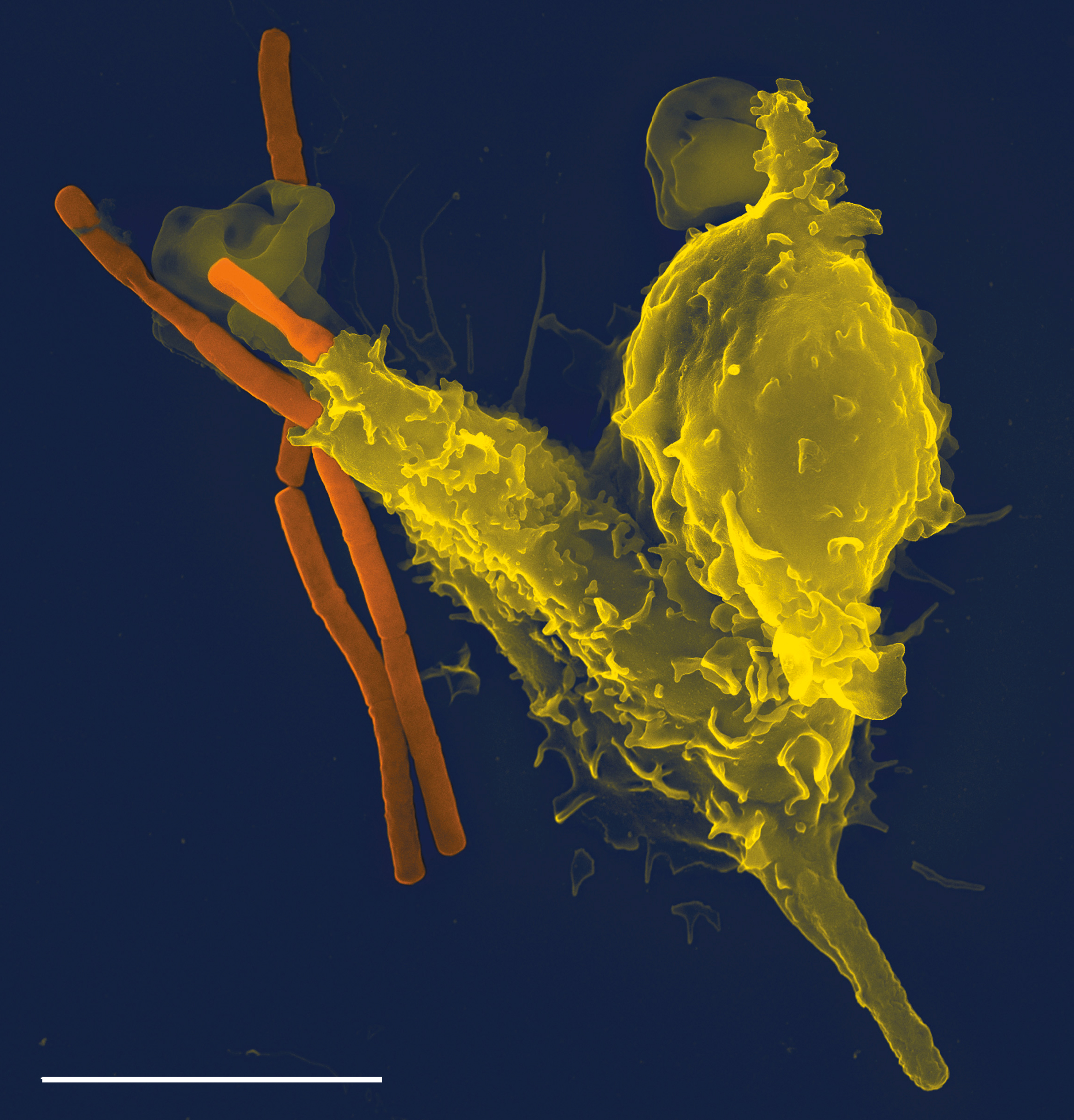
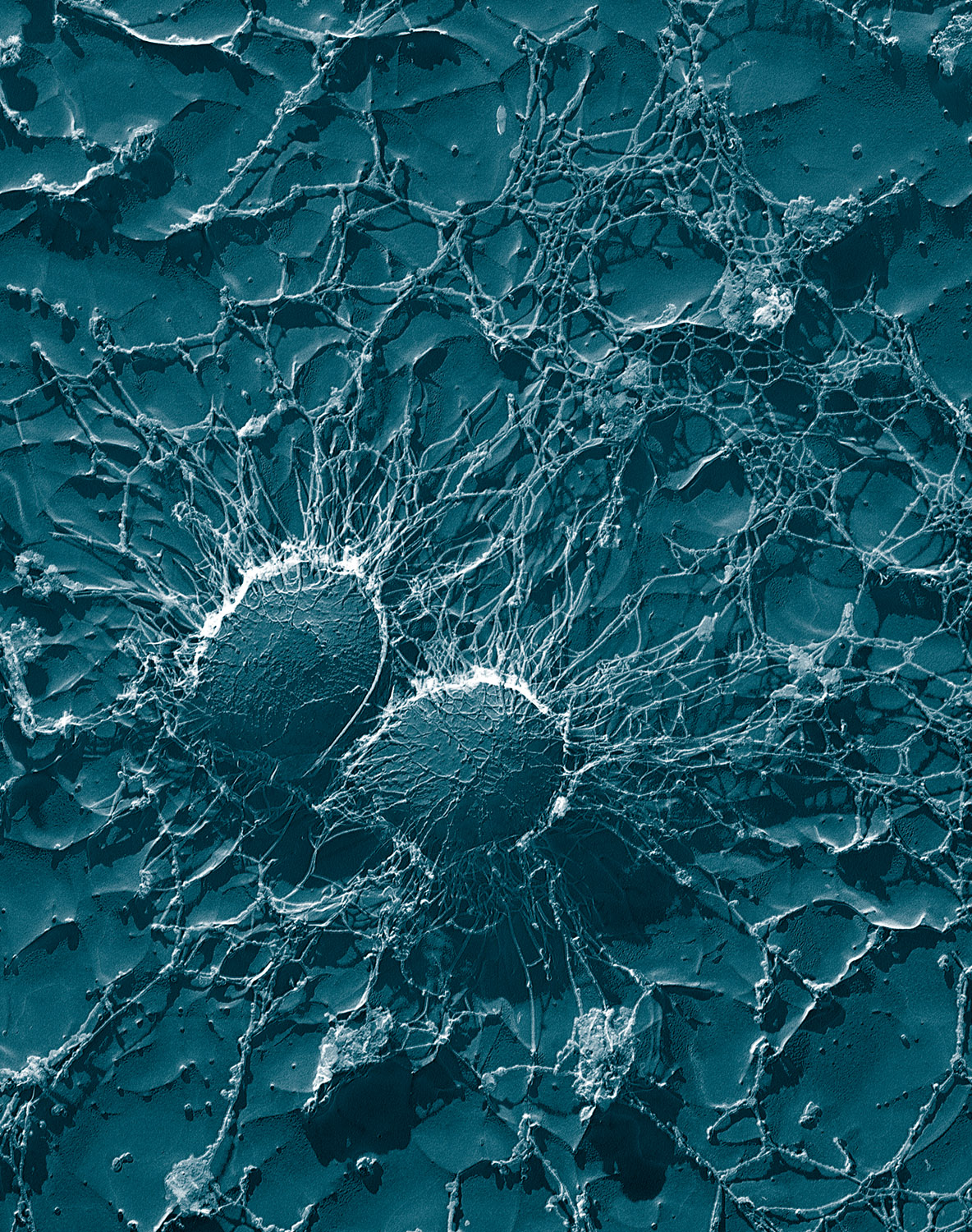
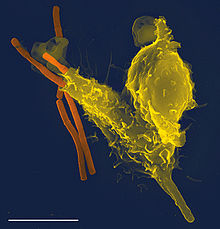 Микрофотография нейтрофила (из электронного микроскопа), фагоцитирующего Bacillus anthracis (оранжевая).
Микрофотография нейтрофила (из электронного микроскопа), фагоцитирующего Bacillus anthracis (оранжевая).
Фагоци́ты — клетки иммунной системы, которые защищают организм путём поглощения (фагоцитоза) вредных чужеродных частиц, бактерий, а также мёртвых или погибающих клеток[1]. Их название произошло от греческого phagein, «есть» или «поедать», и «-cyte», суффикс, в биологии означающий «клетка»[2]. Они важны для борьбы с инфекцией и постинфекционного иммунитета[3]. Фагоцитоз важен для всего животного мира[4] и высоко развит у позвоночных[5]. Фагоциты и фагоцитоз как способ пищеварения у животных были открыты И.И. Мечниковым при изучении губок и плоских червей. Роль фагоцитов в защите от бактерий была впервые открыта И.И.Мечниковым в 1882 году, когда он изучал личинок морских звёзд[6]. Мечников был удостоен в 1908 году Нобелевской премии по физиологии за создание клеточной теории иммунитета[7]. Фагоциты присутствуют в организмах многих видов; некоторые амёбы по многим деталям поведения похожи на макрофаги, что указывает на то, что фагоциты появились на ранних этапах эволюции.[8]
Фагоциты человека и других животных называют «профессиональными» или «непрофессиональными» в зависимости от того, насколько эффективно они фагоцитируют[9]. К профессиональным фагоцитам относятся нейтрофилы, моноциты, макрофаги, дендритные клетки и тучные клетки.[10] Основное отличие профессиональных фагоцитов от непрофессиональных в том, что профессиональные имеют молекулы, называемые рецепторы, на своей поверхности, которые обнаруживают чужеродные объекты, например бактерии.[11] Один литр крови взрослого человека в норме содержит около 2,5—7,5 млрд нейтрофилов, 200—900 млн моноцитов[12].
При инфекции химические сигналы привлекают фагоциты к месту, где патоген проник в организм. Эти сигналы могут исходить от бактерий или от других фагоцитов, уже присутствующих там. Фагоциты перемещаются путём хемотаксиса. Когда фагоциты контактируют с бактериями, рецепторы на их поверхности связываются с ними. Эта связь приводит к поглощению бактерий фагоцитами.[13] Некоторые фагоциты убивают проникших патогенов с помощью оксидантов и оксида азота.[14] После фагоцитоза, макрофаги и дендритные клетки могут также участвовать в презентации антигена — процессе, при котором фагоциты перемещают патогенный материал обратно на свою поверхность. Этот материал затем отображается (презентируется) для других клеток иммунной системы. Некоторые фагоциты поступают в лимфатические узлы и презентируют материал лимфоцитам. Этот процесс важен в формировании иммунитета.[15] Тем не менее, многие болезнетворные микроорганизмы устойчивы к атакам фагоцитов.[3]
История
Русский зоолог Илья Ильич Мечников (1845—1916) впервые определил, что специализированные клетки участвуют в защите от микробной инфекции. В 1882 году, он изучал подвижные клетки в личинках морских звёзд, полагая, что они важны для иммунной защиты животных. Чтобы проверить эту мысль, он воткнул небольшие иголки от мандаринового дерева в личинку. Через несколько часов он обнаружил, что подвижные клетки окружили иголки.[16] Мечников отправился в Вену и поделился своей идеей с Карлом Фридрихом Вильгельмом, который предложил название «фагоцит» для клеток, которые наблюдал Мечников.[17]
Год спустя, Мечников изучал пресноводное ракообразное — Дафнию, маленькое прозрачное животное, которое можно изучать непосредственно под микроскопом. Он обнаружил, что грибные споры, которые нападали на дафний, были уничтожены фагоцитами. Он перенёс свои наблюдения на белые кровяные клетки млекопитающих и обнаружил, что Bacillus anthracis может быть уничтожена фагоцитами. Этот процесс он назвал фагоцитоз.[18] Мечников предположил, что фагоциты — первичная защита от проникающих микроорганизмов.
В 1903 году, Алмрот Райт обнаружил, что фагоцитоз поддерживается специфическими антителами, которые он назвал опсонинами.[19] Мечников был награждён (вместе с Паулем Эрлихом) в 1908 году Нобелевская премия по физиологии или медицине за свою работу по фагоцитам и фагоцитозу.[7]
Хотя значение этих открытий постепенно получило признание в начале двадцатого века, сложные взаимоотношения между фагоцитами и другими компонентами иммунной системы, не были известны вплоть до 1980-х годов.[20]
Фагоцитоз
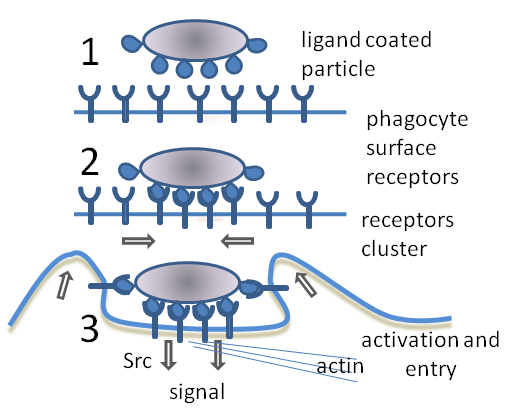
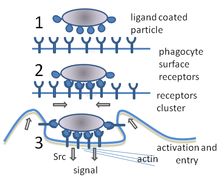 Три стадии фагоцитоза:
Три стадии фагоцитоза:
1. Несвязанные рецепторы на поверхности фагоцита не запускают фагоцитоз.
2. Связывание рецепторов вызывает их группирование.
3. Запускается фагоцитоз и частица поглощается фагоцитом.Фагоцитоз — процесс поглощения чужеродных частиц клетками.[21] Он включает последовательность молекулярных процессов.[22] Фагоцитоз возникает после связывания рецепторами чужеродного агента (например, бактерии). Затем фагоцит окружает бактерию и поглощает её. Фагоцитоз бактерии человеческим нейтрофилом происходит примерно за 9 минут.[23] Внутри фагоцита бактерия оказывается в составе фагосомы. В течение минуты фагосома сливается с лизосомой или гранулой, содержащими ферменты, с образованием фаголизосомы. Заключённая бактерия подвергается агрессивному воздействию[24] и погибает через несколько минут.[23] Дендритные клетки и макрофаги действуют не так быстро и фагоцитоз в этих клетках может протекать в течение многих часов. Макрофаги поглощают большое количество чужеродного материала и часто выделяют некоторые непереваренные частицы обратно. Этот материал является сигналом для миграции макрофагов из крови.[25] Фагоциты способны поглощать почти любое вещество.
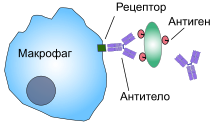 Макрофаги имеют специальные рецепторы, которые способствуют фагоцитозу
Макрофаги имеют специальные рецепторы, которые способствуют фагоцитозуФагоциты имеют на своей поверхности множество различных рецепторов, благодаря которым они связывают чужеродный материал.[3] К ним относятся опсониновые рецепторы, скевенджер-рецепторы и Толл-подобные рецепторы. Опсониновые рецепторы усиливают фагоцитоз бактерий, которые покрыты иммуноглобулином G (IgG) или комплементом. «Комплемент» — комплекс белковых молекул в крови, которые разрушают клетки или помечают их для уничтожения.[26] Скевенджер-рецепторы крупными молекулами поверхности бактериальной клетки и Толл-подобные рецепторы связываются с более специфичными молекулами. Связывание Толл-подобных рецепторов усиливает фагоцитоз и вызывает выброс фагоцитами группы гормонов, которые вызывают воспаление.[3]
Механизмы уничтожения чужеродных агентов
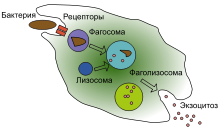 Упрощённая схема фагоцитоза и разрушения бактериальной клетки
Упрощённая схема фагоцитоза и разрушения бактериальной клеткиУничтожение микроорганизмов — важная функция фагоцитоза,[27] которая происходит либо при фагоцитозе (внутриклеточное уничтожение), либо вне фагоцита (внеклеточное уничтожение).
Внутриклеточный кислород-зависимый
Когда фагоцит поглощает бактерию (или любой другой чужеродный материал), увеличивается потребление кислорода, что называют респираторным взрывом. При этом образуются реактивные кислород-содержащие молекулы, которые обладают противомикробным действием.[28] Соединения кислорода токсичны как для патогена, так и для самой клетки, поэтому они хранятся в ячейках внутри самой клетки. Такой метод уничтожения проникающих микроорганизмов называют кислород-зависимое внутриклеточное уничтожение, который делится на 2 типа.[14]
Первый тип — кислород-зависимое образование супероксидного радикала[3], уничтожающего бактерии.[29] Супероксид превращается в пероксид водорода и синглетный кислород под действием фермента супероксиддисмутаза. Супероксиды также взаимодействуют с пероксидом водорода с образованием гидроксильной группы, которая помогает в уничтожении патогенных микробов.[3]
Ко второму типу относят использование фермента миелопероксидаза из нейтрофильных гранул.[30] Когда гранулы сливаются с фагосомой, миелопероксидаза освобождается в фаголизосому, и этот фермент использует пероксид водорода и хлор для создания гипохлорита. Гипохлорит крайне токсичен для бактерий.[3] Миелопероксидаза содержит пигмент гем, за счёт которого образуется зелёный цвет секретов, богатых нейтрофилами (например гной, инфицированная мокрота).[31]
Внутриклеточный кислород-независимый
Фагоциты также могут уничтожать микроорганизмы кислород-независимым методом, но он менее эффективен, чем кислород-зависимый. Различают 4 основных типа. При первом типе используются электрически заряженные белки, которые повреждают клеточную мембрану бактерий. При втором типе используются лизозимы; эти ферменты разрушают клеточную стенку бактерий. При третьем типе используются лактоферрины, которые присутствуют в гранулах нейтрофилов и удаляют необходимое железо из бактерий.[32] При четвёртом типе используются протеазы и гидролазы для переваривания белков разрушенных бактерий.[33]
Внеклеточные
Интерферон-гамма (который также называют фактор, активирующий макрофаги) активирует синтез макрофагами оксида азота. Источником интерферона-гамма могут быть CD4+ T лимфоциты, естественные киллеры, B-лимфоциты, T-киллеры, моноциты, макрофаги или дендритные клетки.[34] Оксид азота затем высвобождается из макрофагов и, в силу своей токсичности, уничтожает микробы вблизи макрофага.[3] Активированные макрофаги образуют и секретируют фактор некроза опухоли. Этот цитокин (класс сигнальной молекулы)[35] уничтожает раковые клетки и клетки, инфицированные вирусом, помогают активировать другие клетки иммунной системы.[36]
При некоторых заболеваниях, например, при редких хронических гранулёматозных заболеваниях, эффективность фагоцитоза нарушается, что может привести к возникновению бактериальных инфекций.[37] При таких заболеваниях существует аномалия в работе различных элементов кислород-зависимого уничтожения микробов. Другие редкие врождённые аномалии, например синдром Шедьяка–Штайнбринка–Хигаси, также связаны с дефектным уничтожением проникающих в организм микробов.[38]
Вирусы
Вирусы могут воспроизводиться только внутри клетки, и они проникают в неё, используя множество рецепторов, участвующих в иммунной защите. Попав внутрь клетки, вирусы используют её биологические процессы в свою пользу, заставляя клетку создавать тысячи вирусных частиц, подобных материнской. Хотя фагоциты и другие компоненты иммунной системы могут в ограниченной степени контролировать вирусы, когда вирус внутри клетки, приобретённый иммунитет (в частности лимфоциты) более важен для защиты.[39] В области вирусной инфекции лимфоцитов скапливается гораздо больше, чем остальных клеток иммунной системы; что наиболее типично для вирусного менингита.[40] Клетки, инфицированные вирусами, уничтожаются лимфоцитами и выводятся из организма фагоцитами.[41]
Роль в апоптозе
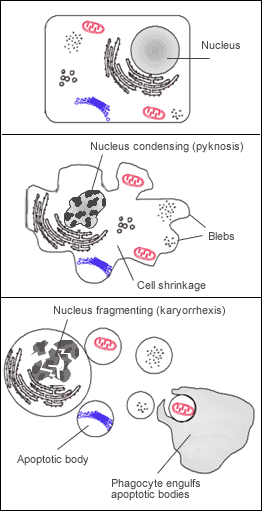
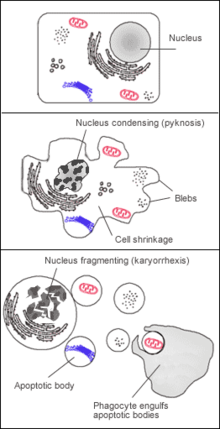 Апоптоз—фагоциты очищают организм от остатков мёртвых клеток
Апоптоз—фагоциты очищают организм от остатков мёртвых клетокУ животных, растений и грибов клетки постоянно погибают. Баланс между делением клетки и их гибелью сохраняет относительно постоянное число клеток у взрослых.[1] Существует 2 механизма гибели клетки: некроз или апоптоз. В отличие от некроза, который чаще возникает как результат заболевания или травмы, апоптоз (или запрограммированная гибель клеток) — нормальный процесс, протекающий в организме. Организм избавляет себя от миллионов мёртвых или погибающих клеток каждый день и фагоциты играют важную роль в этом процессе.[42]
Погибающая клетка, которая подвергается финальной стадии апоптоза,[43] обнажает молекулы, например фосфатидилсерин, на своей поверхности для соединения с фагоцитом.[44] Фосфатидилсерин обычно находится на цитозольной поверхности плазматической мембраны, но перемещается при апоптозе на внешнюю поверхность предположительно с помощью белка под названием скрамблаза.[45] Эти молекулы помечают клетку для фагоцитоза клетками, которые обладают соответствующими рецепторами, например макрофаги.[46] Удаление погибающих клеток фагоцитами происходит упорядоченным образом, не вызывая воспаления.[47]
Взаимодействие с другими клетками
Фагоциты движутся в организме, взаимодействуя с фагоцитарными и нефагоцитарными клетками иммунной системы. Они обмениваются информацией с другими клетками с помощью образования химических веществ, называемых цитокинами, которые вызывают другие фагоциты к области инфекции или активируют «спящие» лимфоциты.[48] Фагоциты составляют часть врождённого иммунитета, который появляется у животных, включая человека, при рождении. Врождённый иммунитет очень эффективен, но не специфичен в отношении определения разницы между видами патогенов. С другой стороны, приобретённый иммунитет более специализированный и может защищать почти от любого вида патогена.[49] Приобретённый иммунитет зависит от лимфоцитов, которые не фагоцитируют, но образуют защитные белки (антитела), которые помечают патогены для уничтожения и предупреждают инфицирование клеток вирусами.[50] Фагоциты, в частности дендритные клетки и макрофаги, стимулируют лимфоциты для образования антител при важном процессе, называемом презентация антигена.[51]
Презентация антигена
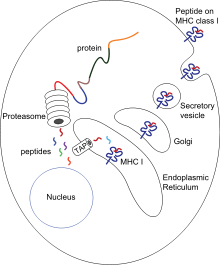 Схема презентации чужеродных пептидов MHC 1 молекулами
Схема презентации чужеродных пептидов MHC 1 молекуламиПрезентация антигена — процесс, при котором некоторые фагоциты перемещают части поглощённого материала назад на свою поверхность и «предоставляют» их для других клеток иммунной системы.[52] Существуют 2 вида «профессиональных» антиген-презентирующих клеток: макрофаги и дендритные клетки.[53] После поглощения, чужеродные белки (антигены) разрушаются до пептидов внутри дендритной клетки или макрофага. Эти пептиды затем связываются с гликопротеинами главного комплекса гистосовместимости (MHC) клетки, которые осуществляют возвращение назад на поверхность фагоциты, где они могут быть «представлены» лимфоцитам.[15] Старые макрофаги не способны быстро перемещаться из области инфицирования, но дендритные клетки могут достигать лимфатических узлов организма, где находятся миллионы лимфоцитов.[54] Это улучшает иммунитет, потому что лимфоциты реагируют на антигены, презентированные дендритными клетками, так же, как если бы они находились в первичной области инфекции.[55] Но дендритные клетки также способны разрушать или подавлять активность лимфоцитов, если они распознают компоненты тела хозяина; это важно для предупреждения аутоиммунных реакций. Этот процесс называют толерантностью.[56]
Иммунологическая толерантность
Дендритные клетки также способствуют иммунологической толерантности,[57], при которой предупреждается иммунологическая атака организма самого себя. Первый тип толерантности — центральная толерантность: когда созревшие T-лимфоциты впервые выходят из тимуса, дендритные клетки разрушают дефектные Т-лимфоциты, которые несут антигены, что может вызывать аутоиммунную реакцию. Второй тип иммунологической толерантности — периферическая толерантность. Некоторые Т-лимфоциты, которые обладают антигенным свойством, способны пройти первый тип толерантности, некоторые Т-лимфоциты развивают аутоиммунные антигены позднее в жизни, и некоторые аутоиммунные антигены не обнаруживаются в тимусе; поэтому дендритные клетки будут работать, опять же, для сдерживания активности аутоиммунных Т-лимфоцитов вне тимуса. Дендритные клетки могут сделать это путём разрушения их или с помощью привлечения Т-супрессоров для дезактивации вредоносных Т-лимфоцитов.[58] Когда иммунологическая толерантность не срабатывает, могут возникнуть аутоиммунные заболевания.[59] С другой стороны, повышенная толерантность может привести к возникновению инфекций, например ВИЧ-инфекции.[58]
Профессиональные фагоциты
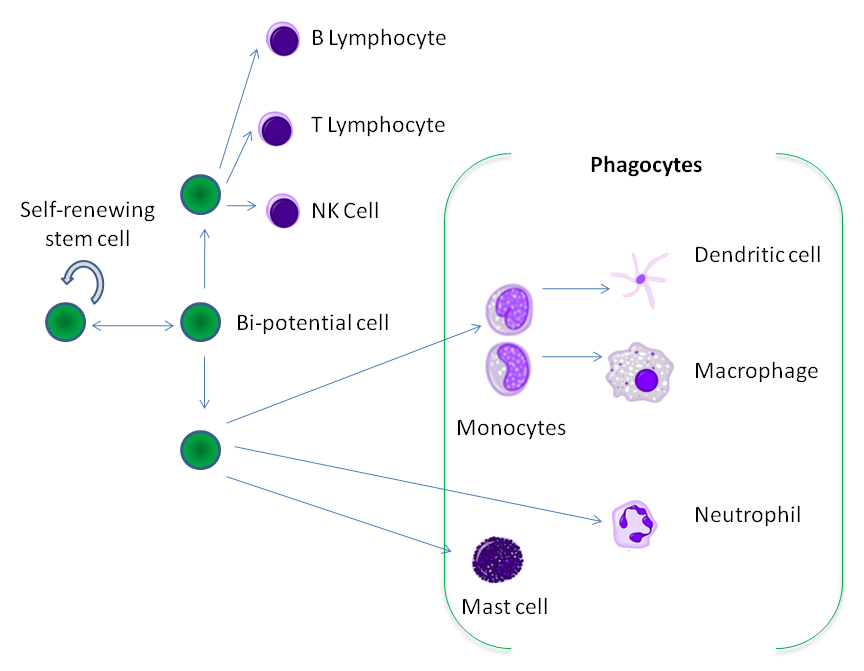
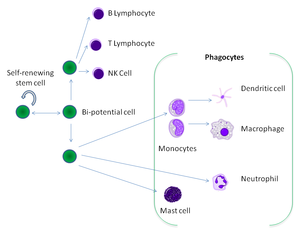 Фагоциты образуются из стволовых клеток костного мозга
Фагоциты образуются из стволовых клеток костного мозгаФагоциты человека и других позвоночных разделяют на «профессиональные» и «непрофессиональные» группы на основе эффективности, при которой они участвуют в фагоцитозе.[9] К профессиональным фагоцитам относят моноциты, макрофаги, нейтрофилы, тканевые дендритические клетки и тучные клетки.[10]
Активация
Все фагоциты, особенно макрофаги, находятся в состоянии готовности. Макрофаги, как правило, относительно пассивны в тканях и размножаются медленно. В таком состоянии полу-покоя они очищают организм от мёртвых клеток и другого неинфекционного мусора и редко принимают участие в презентации антигена. Но при возникновении инфекции они получают химические сигналы (обычно интерферон гамма), которые увеличивают продукцию ими MHC II молекул и подготавливают их к презентации антигена. В таком состоянии, макрофаги — хорошие антиген-презентаторы и киллеры. Однако, если они получают сигнал прямо от патогена, они становятся «гиперактивными», прекращают размножение и концентрируются на уничтожении. Их размер и скорость фагоцитоза увеличивается; некоторые становятся достаточно крупными, чтобы поглотить проникающих в организм простейших.[60]
В крови нейтрофилы неактивны, но движутся по ней с большой скоростью. Когда они получают сигналы от макрофагов из зоны воспаления, они замедляются и выходят из крови. В тканях они активируются цитокинами и поступают в зону действия готовыми уничтожать.[61]
Миграция
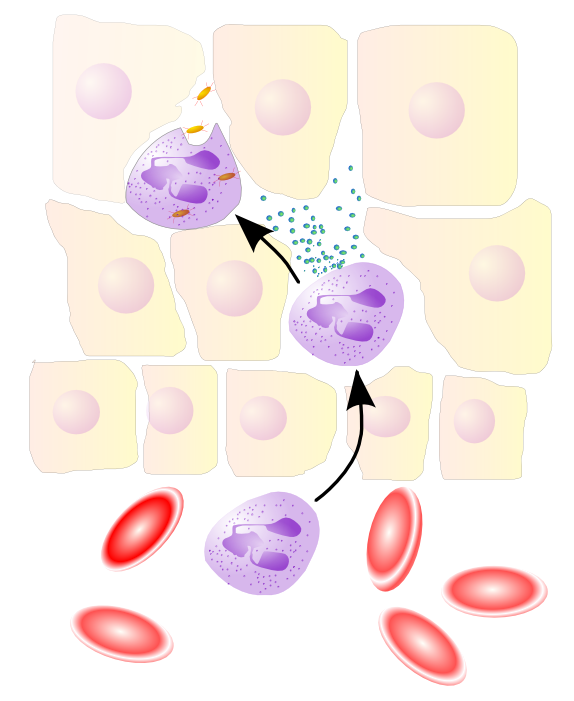
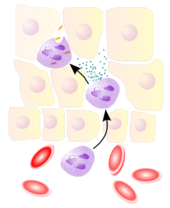 Нейтрофилы выходят из крови в зону инфекции
Нейтрофилы выходят из крови в зону инфекцииКогда происходит инфекционное заражение, химический «SOS» сигнал выделяется для привлечения фагоцитов в зону инфекции.[62] Эти химические сигналы могут включать белки от поступающих бактерий, системы свёртывания пептидов, продукты системы комплемента, а также цитокины, которые выделяются макрофагами, расположенными в ткани в области инфекции.[3] Другая группа химических аттрактантов — цитокины, которые вызывают нейтрофилы и моноциты из кровеносного русла.[13]
Для достижения зоны инфекции, фагоциты выходят из кровеносного русла и проникают в поражённую ткань. Сигналы от инфекции вызывают синтез эндотелиальными клетками, выстилающие кровеносный сосуд, белка, называемого селектин, который сцепляется с проходящими нейтрофилами. Вазодилататоры ослабляют соединительные связи эндотелиальных клеток, что позволяет фагоцитами проходить через стенку сосуда. Хемотаксис — процесс, при котором фагоциты следуют на «запах» цитокинов к области инфекции.[3] Нейтрофилы проникают через органы, покрытые эпителиальной тканью, в зону инфекции, и хотя это важный компонент борьбы с инфекцией, миграция сама по себе может привести к возникновению симптомов заболевания.[63] При инфекции миллионы нейтрофилов вызываются из крови, но они погибают затем в течение нескольких дней.[64]
Моноциты
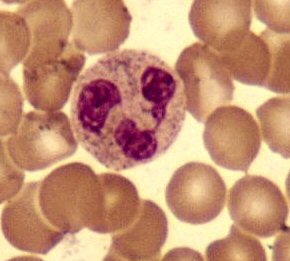
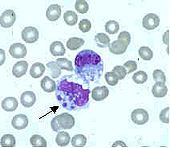 Моноциты с дольчатым ядром в окружении красных кровяных клеток (малое увеличение)
Моноциты с дольчатым ядром в окружении красных кровяных клеток (малое увеличение)Моноциты развиваются в костном мозге и достигают зрелости в крови. Зрелые моноциты имеют крупное, гладкое, дольчатое ядро и цитоплазму, которая содержит гранулы. Моноциты поглощают чужеродные или опасные вещества и презентируют антигены другим клеткам иммунной системы. Моноциты образуют 2 группы: циркулирующая и краевая, которые остаются в других тканях (около 70 % находятся в краевой группе). Большинство моноцитов покидают кровеносное русло через 20-40 часов, попадая в ткани и органы, где они превращаются в макрофаги[65] или дендритические клетки в зависимости от получаемого сигнала.[66] В 1 литре крови человека находится около 500 миллионов моноцитов.[12]
Макрофаги
Зрелые макрофаги не перемещаются быстро, но стоят на страже в тех областях организма, которые подвержены воздействию внешней среды. Там они действуют как сборщики мусора, антиген представляющие клетки или агрессивные киллеры в зависимости от получаемого сигнала.[67] Они образуются из моноцитов, гранулоцитов стволовых клеток или при клеточном делении уже существующих макрофагов.[68] Макрофаги человека диаметром около 21 микрометра.[69]
Этот вид фагоцитов не имеет гранул, но содержит много лизосом. Макрофаги находятся по всему телу почти во всех тканях и органах (например, клетки микроглии в головном мозге и альвеолярные макрофаги в лёгких). Расположение макрофага можно определить по его размеру и внешнему виду. Макрофаги вызывают воспаление путём образования интерлейкина 1, интерлейкина 6 и фактора некроза опухоли.[70] Макрофаги обычно находятся только в тканях и редко попадают в кровоток. Продолжительность жизни тканевых макрофагов, по разным оценкам, от 4 до 5 дней.[71]
Макрофаги могут быть активированы для выполнения функций, которые покоящийся моноцит не может.[70] Т-хелперы подгруппа лимфоцитов, отвечающих за активацию макрофагов. Они активируют макрофаги, посылая сигнал в виде интерферона гамма и экспрессируя белок CD154.[72] Другие сигналы поступают от бактерий в виде фактора некроза опухоли альфа и липополисахаридов.[70] Т-хелперы способны привлекать другие фагоциты в зону инфекции несколькими путями. Они выделяют цитокины, которые действуют на костный мозг, стимулируя образование моноцитов и нейтрофилов и они выделяют некоторые цитокины, которые отвечают за миграцию моноцитов и нейтрофилов в кровеносное русло.[73] Т-хелперы появляются при дифференцировке CD4+ Т лимфоцитов, когда они реагируют на действие антигена в периферических лимфатических тканях.[70] Активированные макрофаги играют важную роль в разрушении опухолей путём образования фактора некроза опухоли альфа, гамма-интерферона, оксида азота, реактивных форм кислорода, катионных белков и гидролитических ферментов.[70]
Нейтрофилы
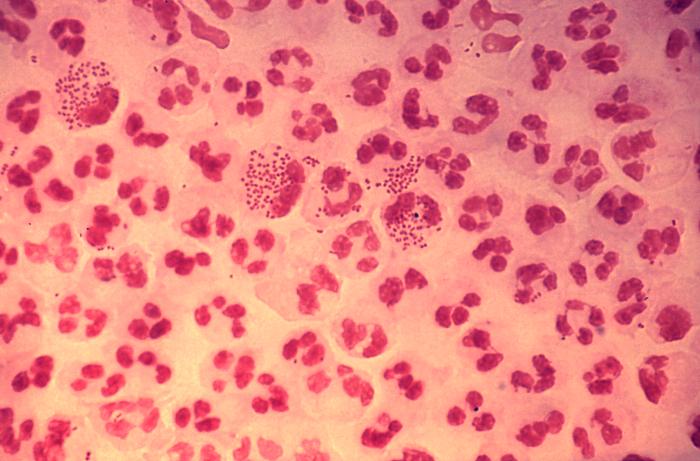
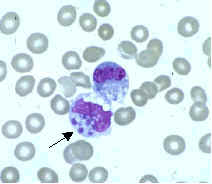
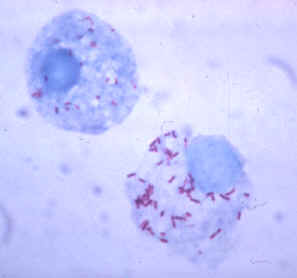
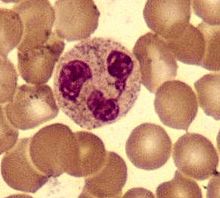 Сегментоядерный нейтрофил (в центре) в окружении эритроцитов, в цитоплазме видны внутриклеточные гранулы (микрофотография, окраска по Романовскому — Гимзе)
Сегментоядерный нейтрофил (в центре) в окружении эритроцитов, в цитоплазме видны внутриклеточные гранулы (микрофотография, окраска по Романовскому — Гимзе)Нейтрофилы обычно находятся в кровеносном русле и являются наиболее распространённым типом фагоцитов, составляя 50-60 % от всех циркулирующих в крови белых кровяных клеток.[74] Один литр крови взрослого человека в норме содержит около 2,5—7,5 миллиардов нейтрофилов.[12] Их диаметр около 10 микрометров[75] и живут только в течение 5 дней.[36] Как только поступает соответствующий сигнал, они в течение примерно 30 минут выходят из крови и достигают зоны инфекции.[76] Они способны быстро поглощать чужеродный материал. Нейтрофилы не возвращаются в кровь; они превращаются в клетки гноя и погибают.[76] Зрелые нейтрофилы меньше, чем моноциты, и имеют сегментированные ядра с несколькими секциями; каждая секция соединяется с хроматиновыми нитями (нейтрофил может иметь 2-5 сегмента). Обычно нейтрофилы не выходят из костного мозга до наступления зрелости, но при инфекции высвобождаются в кровь предшественники нейтрофилов — миелоциты и промиелоциты.[77]
Внутриклеточные гранулы нейтрофилов человека разрушают белки и обладают бактерицидными свойствами.[78] Нейтрофилы способны выделять продукты, которые стимулируют моноциты и макрофаги. Нейтрофильные выделения усиливают фагоцитоз и образование реактивных форм кислорода, участвуя таким образом во внутриклеточном уничтожении.[79] Выделения от первичных гранул нейтрофилов стимулируют фагоциоз бактерий, покрытых IgG.[80]
Дендритные клетки
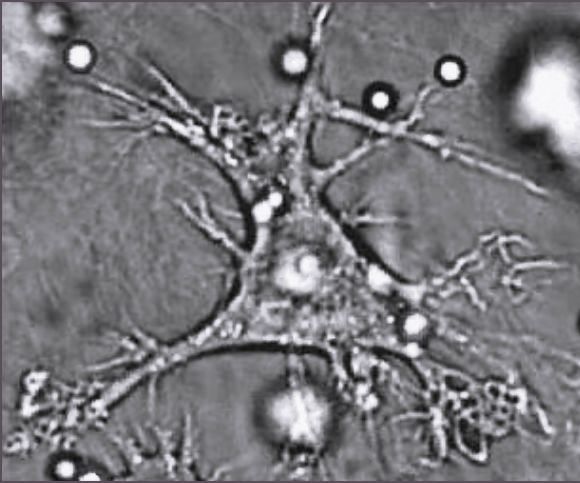
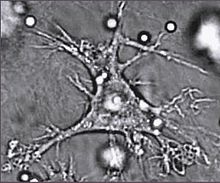 Дендритная клетка
Дендритная клеткаДендритные клетки — специализированные антиген-презентирующие клетки, у которых есть длинные отростки, называемые дендритами,[81] которые помогают поглощать микробы и другие патогены.[82][83] Дендритные клетки находятся в тканях, которые контактируют с окружающей средой, в основном в коже, внутренней оболочке носа, лёгких, желудка и кишечника.[84] После активации, они созревают и мигрируют в лимфатические ткани, где взаимодействуют с Т- и B-лимфоцитами для возникновения и организации приобретённого иммунного ответа.[85] Зрелые дендритные клетки активируют Т-хелперы и Т-киллеры.[86] Активированные Т-хелперы взаимодействуют с макрофагами и B-лимфоцитами чтобы активировать их, в свою очередь. Кроме того, дендритные клетки способны влиять на возникновение того или иного типа иммунного ответа; когда они перемещаются в лимфатические зоны, они способны активировать находящиеся там Т-лимфоциты, которые затем дифференцируют в Т-киллеры и Т-хелперы.[87]
Тучные клетки
Тучные клетки имеют Толл-подобные рецепторы и взаимодействуют с дендритными клетками, Т- и B-лимфоцитами. Тучные клетки выделяют MHC молекулы класса II и могут принимать участие в презентации антигена; однако, роль тучных клеток в презентации антигена ещё не достаточно изучена.[88] Тучные клетки способны поглощать, убивать грамотрицательные бактерии (например, сальмонелла) и обрабатывать их антигены.[89] Они специализируются на обработке фимбриальных белков на поверхности бактерий, которые участвуют в прикреплении к тканям.[90][91] Кроме этих функций, тучные клетки образуют цитокины, которые запускают реакцию воспаления.[92] Это важная часть уничтожения микробов, потому что цитокины привлекают больше фагоцитов к зоне инфекции.[89]
Профессиональные фагоциты[93] Основное расположение Варианты фенотипов Кровь нейтрофилы, моноциты Костный мозг макрофаги, моноциты, синусоидные клетки, обкладочные клетки Костная ткань Остеокласты Кишечник и кишечные бляшки Пейера макрофаги Соединительная ткань Гистиоциты, макрофаги, моноциты, дендритные клетки Печень клетки Купфера, моноциты Лёгкое самовоспроизводящиеся макрофаги, моноциты, тучные клетки, дендритные клетки Лимфатическая ткань свободные и фиксированные и моноциты, дендритные клетки Нервная ткань Клетки микроглии (CD4+) Селезёнка свободные и фиксированные макрофаги, моноциты, синусоидные клетки Тимус свободные и фиксированные макрофаги и моноциты Кожа постоянные клетки Лангерханса, другие дендритные клетки, макрофаги, тучные клетки Непрофессиональные фагоциты
Умирающие клетки и чужеродные организмы поглощаются клетками, отличными от «профессиональных» фагоцитов.[94] К таким клеткам относят эпителиальные, эндотелиальные, паренхиматозные клетки и фибробласты. Их называют непрофессиональными фагоцитами, чтобы подчеркнуть, что в отличие от профессиональных фагоцитов, фагоцитоз для них не основная функция.[95] Фибробласты, например, которые могут фагоцитировать коллаген в процессе ремоделирования шрамов, также способны частично поглощать чужеродные частицы.[96]
Непрофессиональные фагоциты более ограничены, чем профессиональные, в отношении частиц, которые они могут поглотить. Это связано с отсутствием у них эффективных фагоцитарных рецепторов, в частности опсонинов.[11] Кроме того, большинство непрофессиональных фагоцитов не образуют реактивные кислород-содержащие молекулы для фагоцитоза.[97]
Непрофессиональные фагоциты[93] Основное расположение Варианты фенотипов Кровь, лимфа и лимфатические узлы Лимфоциты Кровь, лимфа и лимфатические узлы Естественные киллеры и крупные гранулярные лимфоциты Кожа Эпителиоциты Кровеносные сосуды Эндотелиоциты Соединительная ткань Фибробласты Кровь Эритроциты Устойчивость патогена
 Клетки бактерии Staphylococcus aureus: крупные, волокнистые капсулы, защищающие от атаки фагоцитов.
Клетки бактерии Staphylococcus aureus: крупные, волокнистые капсулы, защищающие от атаки фагоцитов.Патоген вызывает инфекцию, если только он преодолел защиту макроорганизма. Патогенные бактерии и простейшие развивают различные механизмы устойчивости к атакам фагоцитов, и многие из них действительно выживают и размножаются внутри фагоцитирующих клеток.[98][99]
Избежание контакта
У бактерий есть несколько способов избежать контакта с фагоцитами. Во-первых, они могут расти в местах, куда фагоциты не способны попасть (например, повреждённый кожный покров). Во-вторых, бактерия может подавлять реакцию воспаления; без этой реакции фагоциты не способны реагировать на инфекцию адекватно. В-третьих, некоторые виды бактерий могут замедлять способность фагоцитов перемещаться в зону инфекции, препятствуя хемотаксису.[98] В-четвёртых, некоторые бактерии способны избегать контакта с фагоцитом путём обмана иммунной системы, которая начинает «думать», что бактерия — клетка самого макроорганизма. Бледная трепонема (бактерия, вызывающая сифилис) скрывается от фагоцитов, покрывая свою поверхность фибронектином,[100] который естественно образуется в макроорганизме и играет важную роль в заживлении раны.[101]
Избежание поглощения
Бактерии часто образуют белки или сахара, которые покрывают их клетки и препятствуют фагоцитозу; они входят в состав бактериальной капсулы.[98] Например, K5 капсула и O75 O антиген присутствуют на поверхности Escherichia coli,[102] и экзополисахаридные капсулы Staphylococcus epidermidis.[103] Пневмококк образует несколько типов капсул, которые обеспечивают различные уровни защиты,[104] и стрептококки группы А образуют белки, например M белок и фимбриальные белки, которые блокируют процесс поглощения. Некоторые белки препятствуют опсонин-связанному поглощению; Staphylococcus aureus образует белок А для блокирования рецепторов антител, что снижает эффективность опсонинов.[105]
Выживание внутри фагоцита
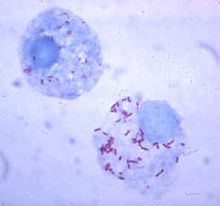 Риккетсия (красным цветом), которая живёт в цитоплазме непрофессионального фагоцита.
Риккетсия (красным цветом), которая живёт в цитоплазме непрофессионального фагоцита.У бактерий развиты способы выживания внутри фагоцитов, где они продолжают уклоняться от иммунной системы.[106] Для безопасного проникновения внутрь фагоцита они выделяют белки, называемые «инвазинами». Оказавшись внутри клетки, они остаются в цитоплазме и избегают воздействия токсических веществ, содержащихся в фаголизосомах.[107] Некоторые бактерии препятствуют слиянию фагосомы и лизосомы.[98] Другие патогены, например лейшмании, образуют высокомодифицированные вакуоли внутри фагоцитов, что позволяет им сохранять устойчивость и размножаться.[108] Legionella pneumophila образует секрет, который заставляет фагоциты сливаться с везикулами, отличными от тех, что содержат токсичные вещества.[109] Другие бактерии способны жить внутри фаголизосом. Staphylococcus aureus, например, образует ферменты каталаза и супероксиддисмутаза, которые разрушают химические вещества (например, пероксид водорода), образуемые фагоцитами для уничтожения бактерий.[110] Бактерии могут выходить из фагосомы перед образованием фаголизосомы: Listeria monocytogenes способна образовывать отверстие в стенке фагосомы, используя ферменты, называемые листериолизин O и фосфолипаза C.[111]
Уничтожение
У бактерий развиты некоторые способы уничтожения фагоцитов.[105] К ним относятся цитолизины, которые образуют поры в клеточной мембране фагоцитов, стрептолизины и лейкоцидины, которые вызывают разрыв гранул нейтрофилов освобождение токсических веществ,[112][113] и экзотоксины, которые снижают поступления АТФ для фагоцитов, необходимых для фагоцитоза. Будучи поглощёнными, бактерии могут убивать фагоциты, высвобождая токсины, которые перемещаются из мембран фагосом или фаголизосом к другим частям клетки.[98]
Нарушение передачи сигнала в клетке
 Множество мелких клеток лейшмании внутри более курпных клеток
Множество мелких клеток лейшмании внутри более курпных клетокНекоторые стратегии выживания часто связаны с нарушением цитокиновой и других методов передачи сигнала в клетке для предотвращения реакции фагоцитов на инвазию.[114] Такие паразитические простейшие как например Toxoplasma gondii, Trypanosoma cruzi и Leishmania инфицируют макрофаги; каждый из них имеет уникальный способ подавления активности макрофагов. Некоторые виды лейшманий изменяют систему сигналов инфицированных макрофагов, подавляют продукцию цитокинов и микробоцидных молекул (оксида азота, реактивных форм кислорода) и нарушают презентацию антигена.[115]
Повреждение макроорганизма фагоцитами
Макрофаги и нейтрофилы, в частности, играют центральную роль в воспалительном процессе путём высвобождения белков и низкомолекулярных воспалительных медиаторов, которые контролируют инфекцию, но могут повреждать ткани макроорганизма. В общем, цель фагоцитов — разрушение патогенов путём поглощения их и последующего воздействия на них токсичными веществами внутри фаголизосом. Если фагоцит не в состоянии поглотить свою цель, эти токсические агенты могут быть выделены в окружающую среду (действие называется «фрустрированный фагоцитоз»). Так как эти агенты токсичны и для клеток макроорганизма, они могут вызвать значительное повреждение здоровых клеток и тканей.[116]
Если нейтрофилы освобождают содержимое гранул (реактивные формы кислорода и протеазы) в почках, то происходит деградация внеклеточного матрикса макроорганизма, что может привести к повреждению клубочковых клеток, влияя на их способность фильтровать кровь и вызывая изменения в их форме. Кроме этого, продукты фосфолипазы (например, лейкотриены) усиливают повреждение. Это освобождение веществ способствует хемотаксису большего количества нейтрофилов в зону инфекции и клубочковые клетки могут быть повреждены ещё сильнее путём прикрепления молекул при миграции нейтрофилов. Повреждения клубочковых клеток может привести к почечной недостаточности.[117]
Нейтрофилы также играют ключевую роль в развитии большинства форм острого повреждения лёгких.[118] При этом активированные нейтрофилы высвобождают содержимое своих токсичных гранул в окружающую среду лёгких.[119] Эксперименты показали, что уменьшение числа нейтрофилов снижают эффекты острого повреждения лёгких,[120] но лечение подавлением нейтрофилов клинически не реалистично, так как при этом организм становится уязвим для инфекций.[119] В печени, повреждение нейтрофилами может способствовать дисфункции и повреждению в ответ на освобождение эндотоксина, выделяемого бактерией, сепсис, травму, алкогольный гепатит, ишемию и гиповолемический шок в результате острой кровопотери.[121]
Химические вещества, высвобождаемые макрофагами, могут также повреждать ткани макроорганизма. Фактор некроза опухоли-α (ФНО) — важное химические вещество, выделяемое макрофагами, которое приводит к свёртыванию крови в мелких сосудах, что предотвращает распространение инфекции.[122] Однако, если бактериальная инфекция проникает в кровь, ФНО-α выделяется в жизненно-важных органах, где может вызвать вазодилатацию и уменьшить объём плазмы; это в свою очередь может привести к септическому шоку. При септическом шоке, высвобождение ФНО-α вызывает блокаду мелких сосудов, которые кровоснабжают жизненноважные органы и может возникнуть их недостаточность. Септический шок может привести к смерти.[13]
Эволюционное происхождение
Фагоцитоз, вероятно, появился на ранних этапах эволюции ,[123] впервые возникнув у одноклеточных эукариот.[124] Амёбы — одноклеточные простейшие, которые отделились от дерева, ведущего к многоклеточным, но они передали множество специфических функций фагоцитарным клеткам млекопитающих.[124] Амёба Dictyostelium discoideum, например, живёт в почве и питается бактериями. Как и фагоциты животных, они поглощают бактерии путём фагоцитоза в основном с помощью Толл-подобных рецепторов и имеют другие биологические функции, общие с макрофагами.[125] Амёбы Dictyostelium discoideum социальны; они слипаются вместе, когда голодают, превращаясь в мигрирующую псевдоплазмодию или личинку. Такой многоклеточный организм в итоге образует плодовое тело со спорами, которое устойчиво к агрессии окружающей среды. Перед образованием плодовых тел, клетки могут перемещаться как личинкоподобные организмы в течение нескольких дней. В течение этого времени воздействие токсинов или бактериальных патогенов может представлять опасность для выживания амёб, ограничивая образование спор. Некоторые амёбы поглощают бактерии и абсорбируют их токсины. В конечном итоге эти амёбы погибают. Они генетически схожи с другими амёбами в личинке и жертвуют собой для защиты других амёб от бактерий, что схоже с самопожертвованием фагоцитов иммунной системы человека. Такая врождённая иммунная функция у социальных амёб предполагает, что древний механизм поглощения клеток мог быть приспособлен для защитной функция задолго до появления разнообразных животных.[126] Но общее происхождение с фагоцитами млекопитающих не доказано. Фагоциты присутствуют во всём животном царстве,[4] начиная от морской губки и заканчивая насекомыми, низшими и высшими позвоночными.[127][128] Способность амёб различать своих и чужих стала основой иммунной системы многих видов.[8]
См. также
Примечания
- ↑ 1 2 Thompson, CB (1995). «Apoptosis in the pathogenesis and treatment of disease». Science 267 (5203): 1456–62. DOI:10.1126/science.7878464. PMID 7878464.
- ↑ The Shorter Oxford English Dictionary. — Oxford University Press (Guild Publishing). — P. 1566–67.
- ↑ 1 2 3 4 5 6 7 8 9 10 Mayer, Gene Immunology — Chapter One: Innate (non-specific) Immunity. Microbiology and Immunology On-Line Textbook. USC School of Medicine (2006). Архивировано из первоисточника 23 августа 2011. Проверено 12 ноября 2008.
- ↑ 1 2 Delves et al. 2006, p. 250
- ↑ Delves et al. 2006, p. 251
- ↑ Ilya Mechnikov, retrieved on November 28, 2008. From Nobel Lectures, Physiology or Medicine 1901—1921, Elsevier Publishing Company, Amsterdam, 1967. Архивная копия от 22 августа 2008 на Wayback Machine
- ↑ 1 2 Schmalstieg, FC; AS Goldman (2008). «Ilya Ilich Metchnikoff (1845–1915) and Paul Ehrlich (1854–1915): the centennial of the 1908 Nobel Prize in Physiology or Medicine». Journal of medical biography 16 (2): 96–103. DOI:10.1258/jmb.2008.008006. PMID 18463079.
- ↑ 1 2 Janeway, Chapter: Evolution of the innate immune system. see Bibliography, retrieved on March 20, 2009
- ↑ 1 2 Ernst & Stendahl 2006, p. 186
- ↑ 1 2 Robinson & Babcock 1998, p. 187 and Ernst & Stendahl 2006, pp. 7–10
- ↑ 1 2 Ernst & Stendahl 2006, p. 10
- ↑ 1 2 3 Hoffbrand, Pettit & Moss 2005, p. 331
- ↑ 1 2 3 Janeway, Chapter: Induced innate responses to infection. see Bibliography, retrieved on March 20, 2009
- ↑ 1 2 Fang FC (October 2004). «Antimicrobial reactive oxygen and nitrogen species: concepts and controversies». Nat. Rev. Microbiol. 2 (10): 820–32. DOI:10.1038/nrmicro1004. PMID 15378046.
- ↑ 1 2 Janeway, Chapter: Antigen Presentation to T Lymphocytes. see Bibliography, retrieved on March 20, 2009
- ↑ Delves et al. 2006, p. 3
- ↑ Aterman K (1 April 1998). «Medals, memoirs—and Metchnikoff». J. Leukoc. Biol. 63 (4): 515–17. PMID 9544583.
- ↑ Ilya Mechnikov. The Nobel Foundation. Архивировано из первоисточника 10 февраля 2012. Проверено 28 ноября 2008.
- ↑ Delves et al. 2006, p. 263
- ↑ Robinson & Babcock 1998, p. vii
- ↑ Ernst & Stendahl 2006, p. 4
- ↑ Ernst & Stendahl 2006, p. 78
- ↑ 1 2 Hampton MB, Vissers MC, Winterbourn CC (February 1994). «A single assay for measuring the rates of phagocytosis and bacterial killing by neutrophils». J. Leukoc. Biol. 55 (2): 147–52. PMID 8301210.
- ↑ Delves et al. 2006, pp. 6–7
- ↑ Sompayrac 2008, p. 3
- ↑ Sompayrac 2008, pp. 13–16
- ↑ Dale DC, Boxer L, Liles WC (August 2008). «The phagocytes: neutrophils and monocytes». Blood 112 (4): 935–45. DOI:10.1182/blood-2007-12-077917. PMID 18684880.
- ↑ Dahlgren, C; A Karlsson (December 17, 1999). «Respiratory burst in human neutrophils.». Journal of Immunological Methods 232 (1–2): 3–14. DOI:10.1016/S0022-1759(99)00146-5. PMID 10618505.
- ↑ Shatwell, KP; AW Segal (1996). «NADPH oxidase.». The international journal of biochemistry and cell biology. 28 (11): 1191–95. DOI:10.1016/S1357-2725(96)00084-2. PMID 9022278.
- ↑ Klebanoff SJ (1999). «Myeloperoxidase». Proc. Assoc. Am. Physicians 111 (5): 383–89. PMID 10519157.
- ↑ Meyer KC (September 2004). «Neutrophils, myeloperoxidase, and bronchiectasis in cystic fibrosis: green is not good». J. Lab. Clin. Med. 144 (3): 124–26. DOI:10.1016/j.lab.2004.05.014. PMID 15478278.
- ↑ Hoffbrand, Pettit & Moss 2005, p. 118
- ↑ Delves et al. 2006, pp. 6–10
- ↑ Schroder K, Hertzog PJ, Ravasi T, Hume DA (February 2004). «Interferon-gamma: an overview of signals, mechanisms and functions». J. Leukoc. Biol. 75 (2): 163–89. DOI:10.1189/jlb.0603252. PMID 14525967.
- ↑ Delves et al. 2006, p. 188
- ↑ 1 2 Sompayrac 2008, p. 17
- ↑ Lipu HN, Ahmed TA, Ali S, Ahmed D, Waqar MA (September 2008). «Chronic granulomatous disease». J Pak Med Assoc 58 (9): 516–18. PMID 18846805. Проверено February 20, 2009.
- ↑ Kaplan J, De Domenico I, Ward DM (January 2008). «Chediak-Higashi syndrome». Curr. Opin. Hematol. 15 (1): 22–29. DOI:10.1097/MOH.0b013e3282f2bcce. PMID 18043242. Проверено April 11, 2009.
- ↑ Sompayrac 2008, p. 7
- ↑ de Almeida SM, Nogueira MB, Raboni SM, Vidal LR (October 2007). «Laboratorial diagnosis of lymphocytic meningitis». Braz J Infect Dis 11 (5): 489–95. PMID 17962876. Проверено March 29, 2009.
- ↑ Sompayrac 2008, p. 22
- ↑ Sompayrac 2008, p. 63
- ↑ Apoptosis. Merriam-Webster Online Dictionary. Архивировано из первоисточника 10 февраля 2012. Проверено 19 марта 2009.
- ↑ Li MO, Sarkisian MR, Mehal WZ, Rakic P, Flavell RA (November 2003). «Phosphatidylserine receptor is required for clearance of apoptotic cells». Science (journal) 302 (5650): 1560–63. DOI:10.1126/science.1087621. PMID 14645847. (Free registration required for online access)
- ↑ Wang X, et al. (2003). «Cell corpse engulfment mediated by C. elegans phosphatidylserine receptor through CED-5 and CED-12». Science 302 (5650): 1563–1566. DOI:10.1126/science.1087641. PMID 14645848. (Free registration required for online access)
- ↑ Savill J, Gregory C, Haslett C. (2003). «Eat me or die». Science 302 (5650): 1516–17. DOI:10.1126/science.1092533. PMID 14645835.
- ↑ Zhou Z, Yu X (October 2008). «Phagosome maturation during the removal of apoptotic cells: receptors lead the way». Trends Cell Biol. 18 (10): 474–85. DOI:10.1016/j.tcb.2008.08.002. PMID 18774293.
- ↑ Sompayrac 2008, p. 44
- ↑ Sompayrac 2008, p. 4
- ↑ Sompayrac 2008, pp. 24–35
- ↑ Delves et al. 2006, pp. 171–184
- ↑ Delves et al. 2006, pp. 456
- ↑ Timothy Lee Antigen Presenting Cells (APC). Immunology for 1st Year Medical Students. Dalhousie University (2004). Архивировано из первоисточника 12 января 2008. Проверено 12 ноября 2008.
- ↑ Delves et al. 2006, p. 161
- ↑ Sompayrac 2008, p. 8
- ↑ Delves et al. 2006, pp. 237–242
- ↑ Lange C, Dürr M, Doster H, Melms A, Bischof F (2007). «Dendritic cell-regulatory T-cell interactions control self-directed immunity». Immunol. Cell Biol. 85 (8): 575–81. DOI:10.1038/sj.icb.7100088. PMID 17592494. Проверено March 29, 2009.
- ↑ 1 2 Steinman, Ralph M. Dendritic Cells and Immune Tolerance. The Rockefeller University (2004). Архивировано из первоисточника 10 февраля 2012. Проверено 15 февраля 2009.
- ↑ Romagnani, S (2006). «Immunological tolerance and autoimmunity.». Internal and emergency medicine. 1 (3): 187–96. DOI:10.1007/BF02934736. PMID 17120464.
- ↑ Sompayrac 2008, pp. 16–17
- ↑ Sompayrac 2008, pp. 18–19
- ↑ Delves et al. 2006, p. 6
- ↑ Zen K, Parkos CA (October 2003). «Leukocyte-epithelial interactions». Curr. Opin. Cell Biol. 15 (5): 557–64. DOI:10.1016/S0955-0674(03)00103-0. PMID 14519390. Проверено March 29, 2009.
- ↑ Sompayrac 2008, p. 79
- ↑ Hoffbrand, Pettit & Moss 2005, p. 117
- ↑ Delves et al. 2006, pp. 1–6
- ↑ Sompayrac 2008, p. 45
- ↑ Takahashi K, Naito M, Takeya M (July 1996). «Development and heterogeneity of macrophages and their related cells through their differentiation pathways». Pathol. Int. 46 (7): 473–85. DOI:10.1111/j.1440-1827.1996.tb03641.x. PMID 8870002.
- ↑ Krombach F, Münzing S, Allmeling AM, Gerlach JT, Behr J, Dörger M (September 1997). «Cell size of alveolar macrophages: an interspecies comparison». Environ. Health Perspect. 105 Suppl 5: 1261–63. DOI:10.2307/3433544. PMID 9400735.
- ↑ 1 2 3 4 5 Bowers, William Immunology -Chapter Thirteen: Immunoregulation. Microbiology and Immunology On-Line Textbook. USC School of Medicine (2006). Архивировано из первоисточника 23 августа 2011. Проверено 14 ноября 2008.
- ↑ Ernst & Stendahl 2006, p. 8
- ↑ Delves et al. 2006, p. 156
- ↑ Delves et al. 2006, p. 187
- ↑ Stvrtinová Viera Neutrophils, central cells in acute inflammation // Inflammation and Fever from Pathophysiology: Principles of Disease. — Computing Centre, Slovak Academy of Sciences: Academic Electronic Press, 1995. — ISBN 80-967366-1-2
- ↑ Delves et al. 2006, p. 4
- ↑ 1 2 Sompayrac 2008, p. 18
- ↑ Linderkamp O, Ruef P, Brenner B, Gulbins E, Lang F (December 1998). «Passive deformability of mature, immature, and active neutrophils in healthy and septicemic neonates». Pediatr. Res. 44 (6): 946–50. DOI:10.1203/00006450-199812000-00021. PMID 9853933. Проверено April 6, 2009.
- ↑ Paoletti, Notario & Ricevuti 1997, p. 62
- ↑ Soehnlein O, Kenne E, Rotzius P, Eriksson EE, Lindbom L (January 2008). «Neutrophil secretion products regulate anti-bacterial activity in monocytes and macrophages». Clin. Exp. Immunol. 151 (1): 139–45. DOI:10.1111/j.1365-2249.2007.03532.x. PMID 17991288. Проверено March 29, 2009.
- ↑ Soehnlein O, Kai-Larsen Y, Frithiof R, et al (October 2008). «Neutrophil primary granule proteins HBP and HNP1-3 boost bacterial phagocytosis by human and murine macrophages». J. Clin. Invest. 118 (10): 3491–502. DOI:10.1172/JCI35740. PMID 18787642. Проверено March 29, 2009.
- ↑ Steinman RM, Cohn ZA (1973). «Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution». J. Exp. Med. 137 (5): 1142–62. DOI:10.1084/jem.137.5.1142. PMID 4573839.
- ↑ Steinman, Ralph Dendritic Cells. The Rockefeller University. Архивировано из первоисточника 10 февраля 2012. Проверено 14 ноября 2008.
- ↑ Guermonprez P, Valladeau J, Zitvogel L, Théry C, Amigorena S (2002). «Antigen presentation and T cell stimulation by dendritic cells». Annu. Rev. Immunol. 20: 621–67. DOI:10.1146/annurev.immunol.20.100301.064828. PMID 11861614. Проверено March 29, 2009.
- ↑ Hoffbrand, Pettit & Moss 2005, p. 134
- ↑ Sallusto F, Lanzavecchia A (2002). «The instructive role of dendritic cells on T-cell responses». Arthritis Res. 4 Suppl 3: S127–32. DOI:10.1186/ar567. PMID 12110131.
- ↑ Sompayrac 2008, pp. 42–46
- ↑ Steinman, Ralph Dendritic Cells. The Rockefeller University. Архивировано из первоисточника 10 февраля 2012. Проверено 16 ноября 2008.
- ↑ Stelekati E, Orinska Z, Bulfone-Paus S (2007). «Mast cells in allergy: innate instructors of adaptive responses». Immunobiology 212 (6): 505–19. DOI:10.1016/j.imbio.2007.03.012. PMID 17544835. Проверено March 29, 2009.
- ↑ 1 2 Malaviya R, Abraham SN (February 2001). «Mast cell modulation of immune responses to bacteria». Immunol. Rev. 179: 16–24. DOI:10.1034/j.1600-065X.2001.790102.x. PMID 11292019. Проверено March 29, 2009.
- ↑ Connell I, Agace W, Klemm P, Schembri M, Mărild S, Svanborg C (September 1996). «Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract». Proc. Natl. Acad. Sci. U.S.A. 93 (18): 9827–32. DOI:10.1073/pnas.93.18.9827. PMID 8790416.
- ↑ Malaviya R, Twesten NJ, Ross EA, Abraham SN, Pfeifer JD (February 1996). «Mast cells process bacterial Ags through a phagocytic route for class I MHC presentation to T cells». J. Immunol. 156 (4): 1490–96. PMID 8568252. Проверено March 29, 2009.
- ↑ Taylor ML, Metcalfe DD (2001). «Mast cells in allergy and host defense». Allergy Asthma Proc 22 (3): 115–19. DOI:10.2500/108854101778148764. PMID 11424870. Проверено March 29, 2009.
- ↑ 1 2 Paoletti, Notario & Ricevuti 1997, p. 427
- ↑ Birge RB, Ucker DS (July 2008). «Innate apoptotic immunity: the calming touch of death». Cell Death Differ. 15 (7): 1096–1102. DOI:10.1038/cdd.2008.58. PMID 18451871.
- ↑ Couzinet S, Cejas E, Schittny J, Deplazes P, Weber R, Zimmerli S (December 2000). «Phagocytic uptake of Encephalitozoon cuniculi by nonprofessional phagocytes». Infect. Immun. 68 (12): 6939–45. DOI:10.1128/IAI.68.12.6939-6945.2000. PMID 11083817.
- ↑ Segal G, Lee W, Arora PD, McKee M, Downey G, McCulloch CA. Involvement of actin filaments and integrins in the binding step in collagen phagocytosis by human fibroblasts. J Cell Sci. 2001 Jan;114(Pt 1):119-129. PubMed PMID: 11112696
- ↑ Rabinovitch M (March 1995). «Professional and non-professional phagocytes: an introduction». Trends Cell Biol. 5 (3): 85–87. DOI:10.1016/S0962-8924(00)88955-2. PMID 14732160.
- ↑ 1 2 3 4 5 Todar, Kenneth Mechanisms of Bacterial Pathogenicity: Bacterial Defense Against Phagocytes. 2008. Архивировано из первоисточника 10 февраля 2012. Проверено 10 декабря 2008.
- ↑ Alexander J, Satoskar AR, Russell DG (September 1999). «Leishmania species: models of intracellular parasitism». J. Cell. Sci. 112 Pt 18: 2993–3002. PMID 10462516.
- ↑ Celli J, Finlay BB (May 2002). «Bacterial avoidance of phagocytosis». Trends Microbiol. 10 (5): 232–37. DOI:10.1016/S0966-842X(02)02343-0. PMID 11973157.
- ↑ Valenick LV, Hsia HC, Schwarzbauer JE (September 2005). «Fibronectin fragmentation promotes alpha4beta1 integrin-mediated contraction of a fibrin-fibronectin provisional matrix». Experimental cell research 309 (1): 48–55. DOI:10.1016/j.yexcr.2005.05.024. PMID 15992798.
- ↑ Burns SM, Hull SI (August 1999). «Loss of resistance to ingestion and phagocytic killing by O(-) and K(-) mutants of a uropathogenic Escherichia coli O75:K5 strain». Infect. Immun. 67 (8): 3757–62. PMID 10417134. Проверено March 29, 2009.
- ↑ Vuong C, Kocianova S, Voyich JM, et al (December 2004). «A crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence». J. Biol. Chem. 279 (52): 54881–86. DOI:10.1074/jbc.M411374200. PMID 15501828. Проверено March 29, 2009.
- ↑ Melin M, Jarva H, Siira L, Meri S, Käyhty H, Väkeväinen M (February 2009). «Streptococcus pneumoniae capsular serotype 19F is more resistant to C3 deposition and less sensitive to opsonophagocytosis than serotype 6B». Infect. Immun. 77 (2): 676–84. DOI:10.1128/IAI.01186-08. PMID 19047408. Проверено March 29, 2009.
- ↑ 1 2 Foster TJ (December 2005). «Immune evasion by staphylococci». Nat. Rev. Microbiol. 3 (12): 948–58. DOI:10.1038/nrmicro1289. PMID 16322743.
- ↑ Sansonetti P (December 2001). «Phagocytosis of bacterial pathogens: implications in the host response». Semin. Immunol. 13 (6): 381–90. DOI:10.1006/smim.2001.0335. PMID 11708894.
- ↑ Dersch P, Isberg RR (March 1999). «A region of the Yersinia pseudotuberculosis invasin protein enhances integrin-mediated uptake into mammalian cells and promotes self-association». EMBO J. 18 (5): 1199–1213. DOI:10.1093/emboj/18.5.1199. PMID 10064587.
- ↑ Antoine JC, Prina E, Lang T, Courret N (October 1998). «The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages». Trends Microbiol. 6 (10): 392–401. DOI:10.1016/S0966-842X(98)01324-9. PMID 9807783.
- ↑ Masek Katherine S. Eurekah Bioscience Collection: Evasion of Phagosome Lysosome Fusion and Establishment of a Replicative Organelle by the Intracellular Pathogen Legionella pneumophila. — Landes Bioscience.
- ↑ Das D, Saha SS, Bishayi B (July 2008). «Intracellular survival of Staphylococcus aureus: correlating production of catalase and superoxide dismutase with levels of inflammatory cytokines». Inflamm. Res. 57 (7): 340–49. DOI:10.1007/s00011-007-7206-z. PMID 18607538. Проверено March 29, 2009.
- ↑ Hara H, Kawamura I, Nomura T, Tominaga T, Tsuchiya K, Mitsuyama M (August 2007). «Cytolysin-dependent escape of the bacterium from the phagosome is required but not sufficient for induction of the Th1 immune response against Listeria monocytogenes infection: distinct role of Listeriolysin O determined by cytolysin gene replacement». Infect. Immun. 75 (8): 3791–3801. DOI:10.1128/IAI.01779-06. PMID 17517863.
- ↑ Datta V, Myskowski SM, Kwinn LA, Chiem DN, Varki N, Kansal RG, Kotb M, Nizet V (May 2005). «Mutational analysis of the group A streptococcal operon encoding streptolysin S and its virulence role in invasive infection». Mol. Microbiol. 56 (3): 681–95. DOI:10.1111/j.1365-2958.2005.04583.x. PMID 15819624.
- ↑ Iwatsuki K, Yamasaki O, Morizane S, Oono T (June 2006). «Staphylococcal cutaneous infections: invasion, evasion and aggression». J. Dermatol. Sci. 42 (3): 203–14. DOI:10.1016/j.jdermsci.2006.03.011. PMID 16679003.
- ↑ Denkers EY, Butcher BA (January 2005). «Sabotage and exploitation in macrophages parasitized by intracellular protozoans». Trends Parasitol. 21 (1): 35–41. DOI:10.1016/j.pt.2004.10.004. PMID 15639739. Проверено March 29, 2009.
- ↑ Gregory DJ, Olivier M (2005). «Subversion of host cell signalling by the protozoan parasite Leishmania». Parasitology 130 Suppl: S27–35. DOI:10.1017/S0031182005008139. PMID 16281989. Проверено March 29, 2009.
- ↑ Paoletti pp. 426-30
- ↑ Heinzelmann M, Mercer-Jones MA, Passmore JC (August 1999). «Neutrophils and renal failure». Am. J. Kidney Dis. 34 (2): 384–99. DOI:10.1016/S0272-6386(99)70375-6. PMID 10430993.
- ↑ Lee WL, Downey GP (February 2001). «Neutrophil activation and acute lung injury». Curr Opin Crit Care 7 (1): 1–7. DOI:10.1097/00075198-200102000-00001. PMID 11373504.
- ↑ 1 2 Moraes TJ, Zurawska JH, Downey GP (January 2006). «Neutrophil granule contents in the pathogenesis of lung injury». Curr. Opin. Hematol. 13 (1): 21–27. DOI:10.1097/01.moh.0000190113.31027.d5. PMID 16319683.
- ↑ Abraham E (April 2003). «Neutrophils and acute lung injury». Crit. Care Med. 31 (4 Suppl): S195–99. DOI:10.1097/01.CCM.0000057843.47705.E8. PMID 12682440.
- ↑ Ricevuti G (December 1997). «Host tissue damage by phagocytes». Ann. N. Y. Acad. Sci. 832: 426–48. DOI:10.1111/j.1749-6632.1997.tb46269.x. PMID 9704069.
- ↑ Charley B, Riffault S, Van Reeth K (October 2006). «Porcine innate and adaptative immune responses to influenza and coronavirus infections». Ann. N. Y. Acad. Sci. 1081: 130–36. DOI:10.1196/annals.1373.014. PMID 17135502. Проверено March 31, 2009.
- ↑ Sompayrac 2008, p. 1
- ↑ 1 2 Cosson P, Soldati T (June 2008). «Eat, kill or die: when amoeba meets bacteria». Curr. Opin. Microbiol. 11 (3): 271–76. DOI:10.1016/j.mib.2008.05.005. PMID 18550419. Проверено April 5, 2009.
- ↑ Bozzaro S, Bucci C, Steinert M (2008). «Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages». Int Rev Cell Mol Biol 271: 253–300. DOI:10.1016/S1937-6448(08)01206-9. PMID 19081545. Проверено April 5, 2009.
- ↑ Chen G, Zhuchenko O, Kuspa A (August 2007). «Immune-like phagocyte activity in the social amoeba». Science (journal) 317 (5838): 678–81. DOI:10.1126/science.1143991. PMID 17673666.
- ↑ Delves et al. 2006, pp. 251–252
- ↑ Hanington PC, Tam J, Katzenback BA, Hitchen SJ, Barreda DR, Belosevic M (April 2009). «Development of macrophages of cyprinid fish». Dev. Comp. Immunol. 33 (4): 411–29. DOI:10.1016/j.dci.2008.11.004. PMID 19063916. Проверено April 5, 2009.
Литература
- Delves P. J. Roitt's Essential Immunology. — 11th. — Malden, MA: Blackwell Publishing, 2006. — ISBN 1405136030
- Phagocytosis of Bacteria and Bacterial Pathogenicity. — New York: Cambridge University Press, 2006. — ISBN 0521845696 Website
- Hoffbrand A. V. Essential Haematology. — 4th. — London: Blackwell Science, 2005. — ISBN 0632051531
- Janeway C. A. Immunobiology. — 5th. — New York: Garland Science, 2001. — ISBN 081533642X
- Phagocytes: Biology, Physiology, Pathology, and Pharmacotherapeutics. — New York: The New York Academy of Sciences, 1997. — ISBN 1573311022
- Phagocyte Function — A guide for research and clinical evaluation. — New York: Wiley–Liss, 1998. — ISBN 0471123641
- Sompayrac L. How the Immune System Works. — 3rd. — Malden, MA: Blackwell Publishing, 2008. — ISBN 9781405162210
При написании этой статьи использовался материал из Энциклопедического словаря Брокгауза и Ефрона (1890—1907).Категории:- Кровь
- Физиология
- Клетки



Wikimedia Foundation. 2010.